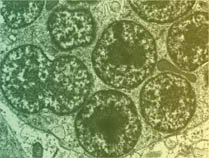
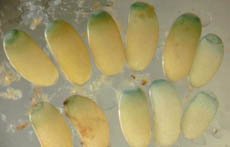
Feature A More Perfect Union Genetic studies show how insects and bacteria within them have teamed up By Jessa Netting May 14, 2001 at 11:28 am - More than 2 years ago Share this:Share Share via email (Opens in new window) Email Share on Facebook (Opens in new window) Facebook Share on Reddit (Opens in new window) Reddit Share on X (Opens in new window) X Print (Opens in new window) Print Their domesticated bacteria enable pea aphids to survive on nutrient-poor plant sap. Charles Hedgecock These Buchnera bacteria live in special cells, called bacteriocytes, inside their aphid hosts. Josh White Aphid eggs come packed with starter populations of their endosymbiotic bacteria (green). Alex Mira Special cells (A) housing this mother aphids helpful bacteria stain bright orange.